कोशिकातत्व
(Cytology)
प्रोटोज़ोआ
(Protoza)
बैक्टीरिया और
वाइरस (Virus)
से ऊँची श्रेणी
के प्रत्येक जंतु
अथवा वनस्पति का
शरीर छोटी
कोशिकाओं से
मिलकर बना होता
है। कोशिकाएँ
इतनी छोटी होती
हैं कि सूक्ष्मदर्शी
के बिना देखी
नहीं जा सकतीं।
प्राणी जितना
ही बड़ा होता
है, वह उतनी ही
अधिक कोशिकाओं
से बना होता
है। जंतुओं और
वनस्पति की कोशिकाओं
में कुछ अंतर अवश्य
होता है, परंतु
साधारण्त: उनकी
संरचना एक ही
ढंग की होती
है। भिन्न भिन्न प्राणियों
की कोशिकाओं
में भी अंतर होता
है। एक ही प्राणी
के विभिन्न अंगों
की कोशिकाओं
के आकार और
गुणों में भी विशेषताएँ
होती हैं, जैसे
किसी भी स्तनधारी
(mammal)
के यकृत और
गुर्दे की कोशिकाओं
की संरचना एक
समान नहीं होती।
इनके कार्य भी
भिन्न हैं। यह विभिन्नता
होते हुए भी कल्पित
साधारण कोशिका
का वर्णन किया
जा सकता है। कोशिका
दो मुख्य भागों
की बनी होती
हैं : (१) केंद्रक कोशिकाद्रव्य
(Cytoplasm)
और (२) केंद्रक (Nucles)
: वानस्पतिक कोशिकाओं
के चारों ओर
सेल्युलोस की
एक भित्ति होती
हैं, परंतु जंतुओं
में ऐसी भित्ति नहीं
मिलती। कोशिकाद्रव्य
में कुछ अंगक होते
हैं जिनका वर्णन
आगे किया जायगा।
लैंगिक प्रजनन करनेवाला प्रत्येक प्राणी अपना जीवन कोशिका अवस्था से ही आरंभ करता है। कोशिका अंडा होती है और इसके निरंतर विभाजन से बहुत सी कोशिकाएँ उत्पन्न हो जाती हैं। कोशिका विभाजन की क्रिया उस समय तक होती रहती है जब तक प्राणी भली भाँति विकसित नहीं हो जाता।
कोशिका विभाजन के समय केंद्रसूत्र दिखाई पड़ते हैं, किंतु स्थित (resting) केंद्रक में ये प्राय: नहीं दिखाई पड़ते। केंद्रक सब ओर एक आवरण से घिरा होता हैं।कोशिकाद्रव्य एक पॉलिफ़ेज़िक कलिल (Polyphasic colliod) है, परंतु यह साधारण कलिलों से भिन्न होता है क्योंकि यह संगठित (organised) होता है। कोशिकाद्रव्य में कई पदार्थ ऐसे होते हैं जो इसकी संरचना में कोई कार्य नहीं करते, किंतु उनका कोशिका के जीवन में बड़ा महत्व है। कोशिकाविभाजन---कोशिका
के प्रत्येक विभाजन
के पूर्व उसके
केंद्रक का विभाजन
होता है। केंद्रकविभाजन
रीत्यनुसार
होनेवाली सुतथ्य
घटना है, जिसे
कई अवस्थाओं में
विभाजित किया
जा सकता है। ये
अवस्थाएँ निम्नलिखित
हैं : (१) पूर्वावस्था
(Prophase),
(२) मध्यावस्था (Metaphase),
(३) पश्चावस्था (Anaphase)
तथा (४) अंत्यावस्था
(Telophase)।
पूर्वावस्था
में केंद्रक के भीतर
पतले पतले सूत्र
दिखाई पड़ते हैं,
जिनको केंद्रकसूत्र
कहते हैं। ये केंद्रकसूत्र
क्रमश: सर्पिलीकरण
(spiralization)
के कारण छोटे
और मोटे हो
जाते हैं। मध्यावस्था
आते समय तक ये
पूर्वावस्था की
अपेक्षा कई गुने
छोटे और मोटे
हो जाते हैं। मध्यावस्था
आने तक कोशिका
के भीतर कुछ
और महत्वपूर्ण
परिवर्तन होते
हैं। केंद्रक का आवरण
नष्ट हो जाता
है और उसकी
जगह एक तर्कुवत्
उपकरण (spindle
apparatus) उत्पन्न होता
है। अधिकांश प्राणियों
की उन कोशिकाओं
में, जिनमें विभाजन
की क्षमता बनी
रहती है, एक विशेष
उपकरण होता
है जिस सेंट्रोसोम
(Centrosomo)
कहते हैं और
जिसके मध्य में
एक कणिका होती
हैं, जिसे ताराकेंद्र
(Centriole)
कहते हैं।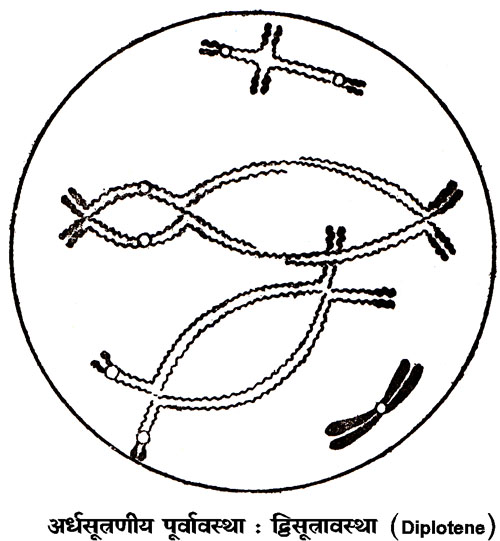
पूर्वावस्था
में ही ताराकेंद्र
का विभाजन हो
जाता है और
एक से दो ताराकें
द्र एक दूसरे को
प्रतिकर्षित (repel)
करते हैं।इसके
कारण ये एक दूसरे
से दूर होते
जाते हैं और
सेंट्रोसोम
दो भागों में
विभाजित हो
जाता है। दोनों
सेंट्रोसोम
एक दूसरे से अधिक
से अधिक दूरी
पर व्यासाभिमुख
(diametrically opposite)
स्थापित हो जाते
हैं। प्रत्येक सेंट्रोसोम
के चारों ओर
कोशिकाद्रव्य
की पतली पतली
रेखाएँ बन जाती
हैं, जिनको ताराकिरण
(Astral rays)
कहते हैं। दोनों
ओर से ताराकिरणें
आकर केंद्रकावरण
पर आघात करती
हैं। इसक समय तक
पूर्वावस्था अपनी
परिसमाप्ति तक
पहुँच जती है
और, जैसा ऊपर
कहा जा चुका
है, केंद्रकावरण
नष्ट हो जाता
है। अब एक सेंट्रोसोम
से लेकर दूसरे
तक तर्कु का प्रसार
होता है। सेंट्रोसोम
और उसकी ताराकिरण
को सेंटर कहते
हैं। तर्कु दो प्रकार
के तर्कुतंतुओं
(Spindle Fibres)
का बना होता
है। एक तो वे तंतु
होते हैं जो
एक सेंटर से दूसरे
सेंटर तक फैले
होते हैं और
जिनको सतत
तंतु (Continuous
Fibres) कहते
हैं। दूसरे वे
तंतु होते हैं,
जिनका एक सिरा
किसी केंद्रकसूत्र
से सटा होता
है और दूसरा
दोनों में से
किसी एक सेंटर
से। मध्यावस्था पर
केंद्रकसूत्र तर्कु
की मध्यरेखा के
समतल पर एकत्रित
हो जाते हैं। इस
समतल को मध्यावस्था
फलक (Metaphase
plate) कहते
हैं। मध्यावस्था में
प्रत्येक केंद्रकसूत्र
अविभाजित ही
प्रतीत होता
है परंतु इसमें
संदेह नहीं कि
इस अवस्था के बहुत
पहले से ही प्रत्येक
केंद्रकसूत्र दो
भागों में विभाजित
रहता है। वस्तुत:
विभाजन की क्रिया
के पूर्व ही अंतराल
अवस्था (Interphase)
में केंद्रक में प्रत्येक
केंद्रकसूत्र अपने
सदृश एक दूसरा
प्रतिवलित 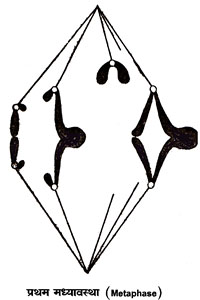 (replicate)
बना लेता है
और ये दोनों
सूत्र एक दूसरे
के इतने समीप
होते हैं कि देखने
में एक ही ज्ञात होते
हैं। प्रत्येक केंद्रकसूत्र
में एक विशेष स्थान
होता है जहाँ
तर्कु का केंद्रकसूत्रीय
तंतु
(chromosomal fibre) जुड़ा
होता है! इसको
सेंटोंमियर
(Centromere) कहते
हैं। किसी किसी
जंतु में केंद्रकसूत्र
मध्यावस्था फलक
के बाह्य भाग में
ही पाए जाते हैं,
परंतु अन्य जंतुओं
में बाह्य भाग और
आंतरिक भाग
दोनों में पाए
जाते हैं। पश्चावस्था
में प्रत्येक केंद्रकसूत्र
के दोनों भाग
एक दूसरे से पृथक्
होने लगते हैं
और इस अवस्था के
अंत काल तक अभिमुखकेंद्र
तक पहुँच जाते
हैं।
(replicate)
बना लेता है
और ये दोनों
सूत्र एक दूसरे
के इतने समीप
होते हैं कि देखने
में एक ही ज्ञात होते
हैं। प्रत्येक केंद्रकसूत्र
में एक विशेष स्थान
होता है जहाँ
तर्कु का केंद्रकसूत्रीय
तंतु
(chromosomal fibre) जुड़ा
होता है! इसको
सेंटोंमियर
(Centromere) कहते
हैं। किसी किसी
जंतु में केंद्रकसूत्र
मध्यावस्था फलक
के बाह्य भाग में
ही पाए जाते हैं,
परंतु अन्य जंतुओं
में बाह्य भाग और
आंतरिक भाग
दोनों में पाए
जाते हैं। पश्चावस्था
में प्रत्येक केंद्रकसूत्र
के दोनों भाग
एक दूसरे से पृथक्
होने लगते हैं
और इस अवस्था के
अंत काल तक अभिमुखकेंद्र
तक पहुँच जाते
हैं।
इसके पश्चात् अंत्यावस्था आरंभ होती है। इस अवस्था में केंद्रकसूत्रों के दोनों समूहों और केंद्रों के चारों ओर केंद्रावरण उतपन्न होते हैं। इस प्रकार एक केंद्रक से दो केंद्रक उत्पन्न होते हैं। जिस समतल पर मध्यावस्था फलक स्थापित था उस स्थान पर एक आवरण बन जाता है, जिसके कारण वह कोशिका दो कोशिकाओं में विभाजित हो जाती है। केंद्रक का विभाजन इसी विधि से होता है। यह बात उपर्युक्त कथन से स्पष्ट है कि इस प्रकार केंद्रकगुणन में और केंद्रकसूत्रों की संख्या में कोई अंतर नहीं होता। केंद्रक विभाजन को समसूत्रण (Mitosis) कहते हैं।
प्राणिजीवन तथा
प्राणिप्रजनन के
हेतु केंद्रक सूत्रों
बड़ा महत्व हैं, क्योंकि
ये आनुवंशिक
पदार्थ से बने
होते हैं। भिन्न
भिन्न जाति के
जंतुओं की कोशिकाओं
में केंद्रकसूत्र
भिन्न भिन्न संख्याओं
में पाए जाते हैं,
परंतु किसी
भी एक जाति के
लिये केंद्रकसूत्रों
की संख्या नियत
होती है और
साधारण अवस्था
में इस संख्या में
कोई विभिन्नता
नहीं होती, जैसे
मनुष्य के शरीर
की प्रत्येक कोशिका
में ४६ केंद्रकसूत्र
होते हैं।
उपरिलिखित
वर्णन साधारण
कोशिकाविभाजन
का है, किंतु कोशिकाविभाजन
का एक विशेष रूप
भी होता है।
अन्य स्थान पर यह
बतलाया गया
है कि जिन प्राणियों
में द्विलैंगिक
प्रजनन की क्रिया
प्रचलित है (और
अधिकांश जंतुओं
में यही क्रिया
पाई जाती है),
उनमें प्राणिजीवन
एक संसेचित अंडे
से आरंभ होता
है। संसेचन में
अंडे के केंद्रक और
शुक्राणु के केंद्रक
का सायुज्य होता
है। सायुज्य का
अर्थ यह हुआ कि
आनुवंशिक पदार्थ
युग्मज में (जो
अंडे और शुक्राणु
(spermatozoon) के
केंद्रक का सायुज्य
होता है सायुज्य
का अर्थ यह हुआ
कि आनुवांशिक
पदार्थ युग्मज
(zygote) में
(जो अंडे और शुक्राणु
के सायुज्य से बनता
है) द्विगुण हो
गया, क्योंकि यह
पदार्थ एक मात्रा
में अंडे में था और
एक मात्रा में शुक्राणु
में। यह स्पष्ट है कि
आनुवंशिक पदार्थ
प्रत्येक पीढ़ी में
द्विगुण नहीं होगा।
संसेचनविधि
में आनुवंशिक
पदार्थ में अनिवार्य
द्विगुणन की क्रिया
इस प्रकार होती
है कि लैंगिक
कोशिकाओं का
परिपक्वताविभाजन
(maturation division)
के समय केंद्रकसूत्रों
की संख्या आधी
हो जाती है। इसका
कारण यह है कि
लैंगिक कोशिकाओं
का परिपक्वताविभाजन,
अथवा अर्धसूत्रण
(Meiosis),
साधारण कोशिकाविभाजन
अथवा समसूत्रण
से भिन्न होता
है। अर्धसूत्रण की
पूर्वावस्था साधारण
समसूत्रण की पूर्वावस्था
की अपेक्षा अधिक
समय तक स्थिर
रहती है और
कई उपावस्थाओं
में विभाजित
की जा सकती हैं।
ये उपावस्थाएँ निम्नलिखित
हैं : (१) लेप्टोटीन
(Leptotene),
(२) ज़ाइगोटीन (Zygotene),
(३) पैकिटीन (Pacytene),
(४) डिप्लोटीन (Diplotene)
तथा (५) डायाकिनीसिस
(Diakinesis)।
इसके पश्चात् मध्यावस्था
और अंत्यावस्था
उसी प्रकार आती
है जैसे साधारण
समसूत्रण में।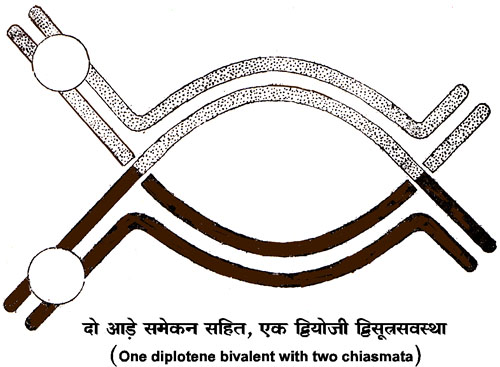
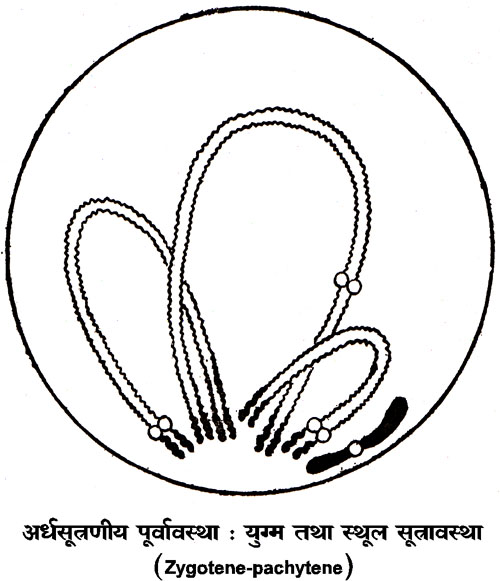
लेप्टोटीन अवस्था में केंद्रक लंबे और पतले केंद्रकसूत्रों से भरा पाया जाता है। इन सूत्रों पर कहीं कही कणिकाएँ पाई जाती हैं, जिनको क्रोमोमियर (Chromomere) कहते हैं। क्रोमोमियरों के बीच के केंद्रकसूत्रों के भागों को इंटर क्रोमोमेरिके फ़ाइब्रिली (interchromomeric fibrillae) कहते हैं। इंट्रोक्रोमोमेरिक फाइब्रिली की अपेक्षा क्रोमोमियर में अभिरंजित होने की अधिक क्षमता होती है। ज़ाइगोटीन उपावस्था में केंद्रकसूत्रों का युग्मन होता है। लैंगिक केंद्रकसूत्रों के अतिरिक्त जीव के केंद्रक में केंद्रकसूत्रों के दो एकात्मक कुलक होते हैं। एक कुलक में कई केंद्रकसूत्र होते हैं, जो साधारणत: एक दूसरे से भिन्न होते हैं। इसका अर्थ यह है कि प्रत्येक प्रकार के दो केंद्रकसूत्र होते हैं। जैसा ऊपर कहा जा चुका है, युग्मांशु उपवस्था में केंद्रकसूत्रों का युग्मन होता है। युग्मन की क्रिया क्रमहीन रूप में नहीं होती, वरन् बहुत क्रमबद्ध होती है। यह क्रिया केवल समान केंद्रकसूत्रों के बीच होती है। प्रत्येक केंद्रकसूत्र अपने समान सूत्र के साथ एक सिरे से दूसरे सिरे तक जुड़ जाता है और जुड़े हुए सूत्रों के क्रोमोमियर केवल अपने समान क्रोमोमियर से ही जुड़ते हैं। ज़ाइगोटीन अवस्था के अंत तक युग्मन की क्रिया पूर्ण हो जाती है। साथी केंद्रकसूत्र एक दूसरे के इतने अधिक समीप होते हैं कि वे एक प्रतीत होते है। केंद्रकसूत्रों के ऐसे जोड़ों को द्विसंयोजक कहा जाता है।
पैकिटीन उपावस्था में प्रत्येक द्विसंयोजक के युग्मित सूत्र एक दूसरे के इतने समीप होते हैं कि पूर्ण द्विसंयोजक देखने में एक सूत्र प्रतीत होता है। पैकिटीन समय में सर्पित संघनन (spiral condensation) के कारण द्विसंयोजक छोटे होने लगते हैं और डिप्लोटीन तथा डायाकिनीसिस समय में द्विसंयोजक और भी छोटे हो जाते हैं। डिप्लोटीन उपावस्था में एक द्विसंयोजक के दोनों सूत्रों में से प्रत्येक सूत्र दो दो सूत्रों में विभाजित हो जाता है। इसका फल यह होता है कि प्रत्येक द्विसंयोजक दो जोड़ी युग्मित सूत्रों से बना पाया जाता है। एक केंद्रकसूत्र के विभाजन से उत्पन्न दो सूत्रों को अर्धसूत्र (Chromatid) कहते हैं। डिप्लोटीन अवस्था में ये युग्मित सूत्र एक दूसरे से पृथक् हो जाते हैं, किंतु कुछ स्थानों पर ये एक दूसरे से अलग नही हो पाते। इसका कारण यह है कि प्रत्येक पक्ष का एक अर्धसूत्र जगह जगह पर टूट जाता है और फिर अभिमुख पक्ष के टूटे हुए एक अर्धसूत्र के दोनों खंडों से इसके खंड जुट जाते हैं। डिप्लोटीन अवस्था में युगल अर्धसूत्र (sister chromatid) एक दूसरे से सटे होते हैं और अभिमुख युगल अर्धसूत्रों से स्पष्टत: दूर होते हैं, परंतु जगह जगह पर उपर्युक्त घटना के कारण एक अर्धसूत्र अपने युगल अर्धसूत्र का साथ छोड़कर अभिमुख पक्ष के अर्धसूत्र के साथ सटा प्रतीत होता हैं। ऐसी संरचनाओं को किऐज़मेटा (Chiasmata) कहते हैं। सूत्रों के अधिक मोटे ओर छोटे होने के कारण डायाकिनिसिस में अर्धसूत्रों का पारस्परिक संबंध सुगमता से नहीं देखा जा सकता और मध्यावस्था (Metaphase) में तो केंद्रकसूत्रों का भूयिष्ठ संघनन हो जाता हैं, जिससे किऐज्म़ेटा की उपस्थित का अनुमान किया जा सकता हैं।
यद्यपि अर्धसूत्रण की पूर्वावस्था (Prophase) से पहले ही प्रत्येक केंद्रकसूत्र का विभाजन हो जाता है, तथापि इनके सेंट्रोमियर का विभाजन मध्यावस्था तक भी नहीं होता। इस कारण विभाजित हो जाने के पर भी प्रत्येक केंद्रकसूत्र की निजता बनी रहती है। पश्चावस्था मे प्रत्येक केंद्रकसूत्र अपने साथी से अलग हो जाता है, अर्थात् प्रत्येक द्विसंयोजक के दोनों केंद्रकसूत्रों का विघटन (dissociation) हो जाता है और युग्मित केंद्रकसूत्रों में से कि सी ध्रुव (Pole) की ओर जाता है और दूसरा उसके विरुद्ध ध्रुव की ओर। इस वर्णन से यह स्पष्ट है कि प्रत्येक ध्रुव पर केंद्रकसूत्र अपनी आधी संख्या में ही पहुँचते हैं।
अंतराल अवस्था बहुत ही अल्पकालीन होती है और कुछ जंतुओं में तो होती ही नहीं। अंतराल अवस्था का अंत होने पर फिर पूर्वावस्था का प्रारंभ होता है। मध्यावस्था, उसके पश्चात् तथा अंत्यावस्था का क्रम वैसा ही होता है जैसा साधारण समसूत्रण में। यह ऊपर कहा जा चुका है कि अर्धसूत्रण में एक के बाद एक, दो बार, कोशिकाविभाजन होता है। इस प्रकार दोनों कोशिकाविभाजन की अवस्थाओं का पृथक् पृथक् निर्दिष्ट करने के लिये उनका मध्यावस्था-१, मध्यावस्था-२, अंत्यावस्था-१, अंत्यावस्था-२ इत्यादि कहते हैं।
यह ऊपर कहा जा चुका है कि पूर्वावस्था से ही प्रत्येक केंद्रकसूत्र दो अर्धसूत्रों में विभाजित हो जाता है, परंतु उसका संट्रोमियर अविभाजित ही रहता है। इसलिये मध्यावस्था-२ पर केंद्रकसूत्र सेंट्रोमियर को छोड़कर पूर्णरूप से विभाजित होता है। पश्चावस्था का प्रारंभ होने पर सेंट्रोमियर दो भागों में विभाजित हो जाता है, जिसके फलस्वरूप केंद्रकसूत्र के दोनों भाग एक दूसरे से मुक्त हो जाते हैं और अभिमुख ध्रुव की ओर जा सकते हैं।
लैंगिक केंद्रकसूत्र विशेष केंद्रकसूत्र होते हैं, जो एक लिंग में युग्मित होते हैं परंतु दूसरे में नही; जैसे ड्रोसॉफ़िला मेलानोगैस्टर (Drosophila melanogaster) में साधारण केंद्रकसूत्रों के तीन जोड़े होते हैं, जिनको आलिंग सूत्र (Autosome) कहते हैं, और दो लैंगिक केंद्रकसूत्र होते हैं। मादा में दोनों लैंगिक केंद्रकसूत्र एक समान होते हैं। इन्हें य-केंद्रकसूत्र (X-chromosome) कहते हैं। नर में भी दो लिंग केंद्रकसूत्र होते हैं। एक य-केंद्रकसूत्र होता है, जो हर मादा य-केंद्रकसूत्र के समान होता है, परंतु दूसरा य-केंद्रकसूत्र से भिन्न होता है। इसे र-केंद्रकसूत्र (Y-chromosome) कहते हैं।
मादा में अर्धसूत्रण के अंत में प्रत्येक कोशिका में चार केंद्रकसूत्र होते हैं-तीन आलिंगसूत्र और एक य-केंद्रकसूत्र। प्रत्येक ऊसाइट (Oocyte) दो बार विभाजित होता है। इससे चार कोशिकाएँ उत्पन्न होती हैं। इनमें से तीन ध्रुवीय पिंड (Polar Bodies) होती है, जिनका शीघ्र ही नाश हो जाता है, और एक परिपक्व अंडाणु (Ovum) होता हैं।
मादा की भाँति नर में प्रत्येक शुक्रकोशिका (Spermatocyte) दो बार विभाजित होती है, जिससे चार स्परमाटिड (Spermatid) उत्पन्न होते हैं।
ये स्परमाटिड दो भाँति के होते है। एक में तीन आलिंग सूत्र और एक य-केंद्रकसूत्र होता है और दूसरे में तीन आलिंग सूत्र और एक र-केंद्रकसूत्र होता है। यह स्पष्ट है कि स्परमाटिड दो प्रकार के होते हैं, परंतु अंडाणु एक ही प्रकार का। प्रत्येक स्परमाटिड क्रमश: लंबा और पतला हो जाता है। इसको स्परमातोज़ोऑन (Spermatozoon) कहते हैं। संसेचन में एक स्परमाटोज़ोऑन का सिर एक अंडाणु में प्रवेश करता है। संसेचित अंडाणु को युग्मज (Zygote) कहते हैं और चूँकि शुक्राणु दो प्रकार के होते है, अत: युग्मज भी दो प्रकार के होते हैं।
एक श्रेणी का युग्मज मादा होता है और दूसरी श्रेणी का नर।
ऐसा भी होता है कि मादा में दो य-केंद्रकसूत्र हों और नर में केवल एक य-केंद्रकसूत्र। ऐसी दशा में लिंगनिर्णय (sex determination) उसी भाँति होता है जैसे भाँति होता है जैसे ड्रोसॉफिला मेलानो-गैस्टर में। नर के शरीर में दो प्रकार के शुक्राणु उत्पन्न होते हैं-एक में आलिंग सूत्र के अतिरिक्त य-केंद्रकसूत्र होता है और दूसरे में य-केंद्रकसूत्र होता ही नहीं। ऐसे भी जंतु हैं जिनके नर में परस्पर भिन्न कई य-केंद्रकसूत्र होते हैं। अर्धसूत्रण के अंत पर दो प्रकार के स्परमाटिड बनते है। एक प्रकार के स्परमाटिड में आलिंगसूत्र के अतिरिक्त य१, य२, य३ इत्यादि केंद्रकसूत्र होते हैं और दूसरे में केवल र-केंद्रकसूत्र और आलिंगसूत्र।
केंद्रकसूत्र की संरचना में दो पदार्थ विशेषत: संमिलित रहते हैं-(१) डिआक्सीरिबोन्यूक्लीइक अम्ल (Deoxryibonucleic acid) तथा (२) हिस्टोन (Histone) नामक एक प्रकार का प्रोटीन। डिआक्सीरिबोन्यूक्लीइक अम्ल डी एन ए (D N A) ही आनुवंशिक (hereditary) पदार्थ है। डी ए न ए (D N A) अणु की संरचना में चार कार्बनिक समाक्षार सम्मिलित होते हैं : दो प्यूरिन (purines), दो पिरिमिडीन (pyrimidines), एक चीनी-डिआक्सीरिबोज़्ा (Deoxyribose) और फासफ़ोरिक अम्ल (Phosphoric acid)। प्यूरिन में ऐडिनिन (Adenine) और ग्वानिन (Guanine) होते है और पिरिमिडीन में थाइमीन (Thymine) और साइटोसिन (Cytosine)। डी एन ए (D N A) के एक अणु में दो सूत्र होते हैं, जो एक दूसरे के चारों और सर्पिल रूप में वलयित (spirallyicoiiled) होते है। प्रत्येक डी एन ए (D N A) सूत्र में एक के पीछे एक चारों कार्बनिक समाक्षार इस क्रम से होते हैं-थाइमीन, साइटोसिन, ऐडिनीन और ग्वानिन, एवं वे परस्पर एक विशेष ढंग से जुड़े होते हैं।
इन चार समाक्षारों और उनसे संबंधित शर्करा और फास्फोरिक अम्ल अणु का एक एकक टेट्रान्यूक्लीओटिड (Tetranucleotide) होता है और कई सहस्त्र टेट्रान्यूक्लीओटिडों का एक डी एन ए (D N A) अणु बनता है।
विभिन्न प्राणियों के डी एन ए की विभिन्नता का कारण समाक्षारों के अनुक्रम में अंतर है। डी एन ए और ऐसा ही एक दूसरा न्यूक्लीइक अम्ल आर एन ए (R N A) कार्बनिक समाक्षार की उपस्थिति के कारण पराबैंगनी को अधिकांश २,६०० आं. के क्षेत्र में अंतर्लीन करते हैं। इसी आधार पर डी एन ए का एक कोशिका संबंधी मात्रात्मक आगमन किया जाता है।
प्राणियों में दो विशेष प्रकार के केंद्रसूत्र पाए जाते हैं। एक तो कुछ डिप्टरा इंसेक्टा (Diptera, Insecta) में डिंभीय लारग्रंथि (larval salivary gland) के केंद्रकों में पाया जाता है। ये केंद्रकसूत्र उसी जाति के साधारण केंद्रकसूत्रों की अपेक्षा कई सौ गुने लंबे और चौड़े होते हैं। इस कारण इन्हें महाकेंद्रकसूत्र (Giant chromosomes) कहते हैं। इनकी संरचना साधारण समसूत्रण और अर्धसूत्रण केंद्रसूत्रों से कुछ भिन्न दिखाई पड़ती है। यहाँ एक केंद्रकसूत्र के स्थान पर एक अनुप्रस्थ पंक्ति ऐसी कणिकाओं की होती है जिनमें अभिरंजित होते की योग्यता अधिक होती हैं। केंद्रकसूत्र के एक छोर से दूसरे तक बहुत सी ऐसी अनुप्रस्थ पंक्ति की सब कणिकाएँ एक समान होती हैं और अन्य पंक्तियों की कणिकाओं में विशेषताएँ और विभिन्नताएँ होती है। इन केंद्रकसूत्रों के अधिक लंबे होने के कारण यह समझा जाता है कि इनका पूर्ण रूप से विसर्पिलीकरण (despiralisation) होता है और कदाचित् प्रोटीन का कुछ बढ़ाव भी होता हैं।
अधिक चौड़े होने के कारण यह है कि एक केंद्रकसूत्र अपने समान एक दूसरे केंद्रक-त्र का संश्लेषण करता है। साधारण अवस्था में समसूत्रण के समय ये दोनों सूत्र एक दूसरे से पृथक् हो जाते हैं, परंतु महाकेंद्रकसूत्र में यह नहीं होता। दोनों सूत्र एक दूसरे से जुड़े ही रह जाते हैं। महाकेंद्रकसूत्र की संख्या साधारण केंद्रकसूत्र की संख्या की आधी होती है, क्योंकि प्रत्येक सूत्र अपने समान दूसरे सूत्र से युग्मित हो जाता है। इस घटना को दैहिक युग्मन (Somatic pairing) कहते हैं।
जंतुओं में विचित्र प्रकार का एक और भी केंद्रकसूत्र पाया जाता है। इसक लैंपब्रश केंद्रकसूत्र (Lampbrush chromosome) कहते हैं। ये केंद्रकसूत्र ऐसे जंतुओं के अंडों के केंद्रकों में पाए जाते हैं जिनमें अंडपीत की मात्रा अधिक होती है, जैसे मछली, उभयचर, उरग, पक्षीगण इत्यादि। केंद्रकसूत्र साधारण डिप्लोटीन-डायाकिनीसिस (Diplotene-diakinesis) केंद्रकसूत्रों के समान दो दो युग्मित सूत्रों के बने होते हैं। दोनों युग्मित सूत्र कुछ स्थानों पर एक दूसरे से जुड़े होते हैं और शेष स्थानों पर एक दूसरें से दूर दूर रहते हैं। इन जोड़ों को किएज्मा समझा जाता है। प्रत्येक सूत्र पर, जिसकों क्रोमोनिमा (Chromonema) कहते हैं, स्थान स्थान पर विभिन्न परिसाण की कणिकाएँ होती हैं जिनको क्रोमिमियर्स (Chromomeres) कहते हैं। प्रत्येक क्रोमोमियर से एक जोड़ी या अधिक पार्श्वपाश (lateral loops) जुड़े हुए होते है। पार्श्वपाश भी क्रोमोनिमा के सदृश सूत्र का बना होता है, परंतु इसके चारों ओर रिबोन्यूक्लिओ-प्रोटीन कणिकाएँ एकत्रित हो जाती हैं जिससे ये सूत्र मोटे दिखाई देते हैं। क्रोमोमियर भी क्रोमोनिमा से संतत होते हैं। केंद्रकाएँ विशेष केंद्रकसूत्र पर उत्पन्न होती हैं।
अधिकांश जंतुओं की प्रत्येक कोशिका में केंद्रकसूत्रों के दो एकात्म कुलक होते हैं। परिपक्व लिंगकोशिकाओं (mature sex-cells) में एक कुलक रह जाता है। ऐसे प्राणी और कोशिकाएँ द्विगणित (diploid) कही जाती हैं, परंतु कुछ प्राणियों, विशेषत: पौधों, में दो से अधिक कुलक केंद्रकसूत्रों के होते हैं यह बहुगुणित (polyploid) कहे जाते हैं।
यदि किसी द्विगुणित प्राणी के केंद्रकसुत्र दुगुने हो जाए, जिससे उसकी कोशिकाएँ में प्रत्येक सूत्र चार चार हों जैसे, (क१ क१ क१ क१; ख१ ख१ ख१ ख१, ग१ ग१ ग१ ग१ इत्यादि) तो ऐसे प्राणी का आटोपॉलिप्लॉइड (आटीटेट्राप्लॉइड) [autopolyploid (autotetraploid)] कहते हैं। यदि किसी द्विगुणित संकर (deploid hybrid) के केंद्रकसूत्र दुगुने हो जाए तो ऐसे प्राणी को ऐलोपॉलिप्लॉइड (allopolyploid) कहते हैं। यदि एक द्विगुणित प्राणी का, जिसकेकेंद्रकसुत्र क१ क१ ख१ ख१ ग१ ग१ इत्यादि हैं, किसी दूसरे प्राणी से, जिसके केंद्रकसूत्र क२ क२ ख२ ख२ ग२ ग२ इत्यादि हैं, संकरण किया जाए तो उसकी संतान के केंद्रकसूत्र क१ क२ ख१ ख२ ग१ ग२ इत्यादि होंगे। क१ क२ ख१ ख२ इत्यादि एक दूसरे से भिन्न होंगे और इनमें साधारणत: युग्मन नहीं होगा। यदि इस प्राणी के केंद्रकसूत्र दुगुने हो जाए तो उनकी कोशिकाएँ में क१ क१ क२ क२; ख१ ख१ख२ ख२; ग१ ग१ ग२ ग२, इत्यादि केंद्रकसूत्र होंगे। यह ऐलोपॉलिप्लॉइड (ऐलोटेट्राप्लॉइड) कहा जाएगा। पॉलिप्लॉइड में चार से अधिक कुलक भी हो सकते हैं।
यह स्पष्ट है कि ऑटोपॉलिप्लॉइड में ज़ाइगोटीन अवस्था में चतु:संयोजक (quadrivalents) उत्पन्न हो जायँगे, क्योंकि प्रत्येक प्रकार के चार चार केंद्रकसूत्र उपस्थित हैं और चार सूत्रों के युग्मन से एक चतु:संयोजक बनता है। कोशिकाविभाजन के समय प्रत्येक ध्रुव को बराबर बराबर संख्या में केंद्रकसूत्र नहीं मिलेंगे। प्राय: ऐसा होता है कि एक चतु:संयोजक के टूटने से किसी ध्रुव पर तीन सूत्र पहुँचे और उसके संमुख ध्रुव पर एक ही सूत्र पहुँचे। कोशिकाविभाजन के अंत पर बने हुए संतति कोशिकाओं (daughter cells) में केंद्रकसूत्र या तो अधिक संख्या में होंगे या कम में और ऐसे असंतुलन का परिणाम यह होता है कि कोशिका मर जाती है। इसी कारण ऑटोटेट्राप्लॉइड बहुत कम उर्वर होते हैं। ऑटोटेट्राप्लॉइड पौधे साधारण द्विगुणित पौधों से बहुत बड़े होते हैं तथा उनके बीज भी बहुत बड़े होते हैं, जिससे उर्वरता कम होने पर भी ये गृहस्थी के लिये अधिक लाभदायक सिद्ध हो सकते हैं। ठंढक पहुँचाकर, या कुछ ऐलकेलायडों के प्रभाव से, पौधे आटोपॉलिप्लाइड बनाए जा सकते हैं।
ऐलोटेट्राप्लॉइड में दशा इसके विपरीत होतीे हैं। यदि दोनों आदिम मातापिता के सूत्र एक दूसरे से पूर्ण रूप से विभिन्न हों तो ऐलोपॉलिप्लाइड क्रियात्मक रूप से द्विगुणित है और पूर्ण रूप से उर्वर होगा। जैसे, यदि किसी संकर में क१ क२,ख१ ख२, ग१ ग२ से सूत्र सर्वथा भिन्न हों तो ऐसा संकर बंध्या होगा, परंतु इसके केंद्रकसूत्रों के दुगुने होने से यह अवस्था बदल जायगी। ऐसी कोशिकाओं में क१ क१ क२ क२, ख१ ख१ ख२ ख२, ग१ ग१ ग२ ग२ इत्यादि सूत्र होंगे और जिन शाखाओं में ऐसी कोशिकाएँ होंगी उनपर फूल लगेंगे, क्योंकि ऐसी कोशिकाओं में माइओटिक विभाजन सफल होगा, क१ क१ से युग्मित होगा, ख१ ख१ से इत्यादि।
धतूरा स्ट्रामोनिअम (Datura stramonium) में द्विगुणित अवस्था में १२ जोड़ी केंद्रकसूत्र होते हैं और अर्धसूत्रण के समय द्विसंयोजक बनते हैं। इसके ऑटोपॉलिप्लॉइड में १२ चतुष्क (४८)केंद्रकसूत्र होते है और अर्धसूत्रण के समय १२ चतु:संयोजक बनते हैं। इसी भाँति प्रिम्यूला साइनेंसिस (Primula sinensis) से द्विगुणित पौधे में १२ जोड़ी केंद्रकसूत्र होते हैं और ऑटोटेट्राप्लॉइड में ४८ सूत्र होते हैं एवं अर्धसूत्रण के समय इसमें ९ से ११ चतु:संयोजक और २ से ६ तक द्विसंयोजक बनते हैं। सोलेमन लाइकोपरसिकॉन (Solanum lycopersicon) के द्विगुणित में १२ जोड़ी केंद्रकसूत्र होते हैं और उसके ऑटोटेट्राप्लॉइड में १२ चतुष्क (४८) केंद्रकसूत्र। ये सब पौधे हैं।
क्रीपिस रुब्रा (Crepis rubra) औरा क्रीपिस फ़ोएटिडा (Crepis foctida) में ५ जोड़ी केंद्रकसूत्र होते हैं। इनके सूत्र एक दूसरे से बहुत भिन्न नहीं होते और इनके संकरण से उत्पन्न संकर में अर्धसुत्रण के समय ५ द्विसंयोजक बनते हैं। इसके ऐलोपॉलिप्लॉइड में २० कें द्रकसूत्र होते हैं और अर्धसूत्रण में ० से ५ चतु:संयोजक बनते हैं और ० से १० द्विसंयोजक। स्पष्ट है कि ऐलोटेट्राप्लॉइड बहुत उर्वर नहीं होगा। प्रिम्यूला फ्लोरिबंडा (Primula floribunda) और प्रिम्यूला रेस्टिसिलेटा (Primula resticillata) दोनों में ही ९ जोड़ी केंद्रकसूत्र होते हैं, जो एक दूसरे के प्राय: समान होते हैं। इनके संकरण से जो संकर बनाता है उसके प्रित्यूजा किवेन्सिस कहते हैं। इसमें भी ९ जोड़ी केंद्रकसूत्र होते हैं। अर्धसूत्रण के समय में युग्मन क्रिया सफल होती है और ९ द्विसंयोजक बनते हैं। सूत्रों के द्विगुण होने से जो ऐलोपॉलिप्लाइड बनता है उसमें ९ चतुष्क (३६) सूत्र होते हैं और ऐसे पौधे में १२ से १८ तक द्विसंयोजक बनते हैं और ० से ३ तक चतु:संयोजक। स्पष्ट है कि चतु:संयोजाकों की संख्या बहुत कम है और कभी कभी एक भी चतु:संयोजक नहीं बनता।
मूली (Raphanus) और करमकल्ला (Brassica) में से प्रत्येक में ९ जोड़ी केंद्रकसूत्र होते हैं, जो एक दूसरे से पूर्णत: भिन्न होते हैं। इनके संकरण से उत्पन्न संकर रैफ़ानस ब्रैसिका (Raphanus-Brassica) में भी ९ जोड़ी केंद्रकसूत्र होते हैं; परंतु अर्धसूत्रण में एक ही द्विसंयोजक नहीं बनता, क्योंकि युग्मन की क्रिया सफल नहीं होती और सभी सूत्र अयुग्मित रह जाते हैं जिससे १२ एक-संयोजक बनते हैं। इसके सूत्र द्विगुण से उत्पन्न ऐलोटेट्राप्लॉइड में १२ जोड़ी सूत्र होते हैं और अर्धसूत्रण में १२ संयोजक बनते हैं, चतु:संयोजक एक भी नहीं। परिणाम यह होता है कि रैफ़ानस-ब्रैसिका-ऐलाटेट्राप्लॉइड बहुत उर्वर होता है, यद्यपि रैफ़ानस-ब्रैसिका-द्विगुणित बंध्या होता है।
जंतुओं में पॉलिप्लॉइडी बहुत कम पाई जाती है पर यह अनिषेकजनित (Parthenogenetic) जंतुओं में बहुधा पाई जाती है। पौधों में बहुत सी नई जातियाँ पॉलिप्लॉइड के कारण उत्पन्न हुई होंगी। इसका प्रमाण इससे मिलता है कि ऐंजिऔस्पर्मों (Angiosperms) की लगभग आधी जातियाँ ऐसी है जिनके परिपक्व युग्मकों (Gametes) के केंद्रकसूत्रों की संख्या किसी संबंधित जाति के युग्मकीय केंद्रकसूत्र की संख्या की गुणित है। गेहूँ की कई जातियाँ हैं। इन जातियों की मूल युग्मकीय केंद्रकसुत्र संख्या ७ है। केंद्रकसूत्रों की संख्या गेहूँ की जातियों में ७ की गुणित १४,२१,४२ तथा ४९ तक पाई जाती है। इसी भाँति तंबाकू की भिन्न भिन्न जातियों में केंद्रकसूत्रों की संख्या १२ अथवा १२ की गुणित २४ होती है। पौधों में प्रयोग द्वारा बहुत से पॉलिप्लॉइड बनाए गए हैं, जिनमें एकात्मक सूत्रों के दो कुलक होते हैं। ये उर्वर होते हैं।
इसमें संदेह नहीं कि कोशिकाद्रव्य (Cytoplasm) केंद्रक के नियंत्रण में कार्यशील होता है। अनेक प्रकार के कोशिकासमूह अन्यान्य कार्यों के संचालन में लगे रहते हैं। उदाहरणत: अग्न्याश (Pancreas) की एक्सोक्राइन (Excorine) कोशिकाएँ विशेष पाचक किण्वज उत्पन्न करती हैं। गुद की नलिका की कोशिकाएँ रुधिर से यूरिया निकाल लेती हैं और यकृत की कोशिकाएँ ग्लूकोस को ग्लाइकोजन में परिणत करके एकमित कर लेती हैं। स्पष्ट है कि किसी भी प्राणी की प्रत्येक कोशिका में उसके सब जीन (Gene) साधारणत: उपस्थित होते हैं। इसलिये भिन्न भिन्न प्रकार की कोशिकाओं के विविध प्रकार के प्रोटीनों (जिनकी वे बनी है) की उत्पत्ति में कुछ उपयुक्त जीन तो सक्रिय रहे होंगे और शेष सब निष्क्रिय हो गए होंगे ओर उनकी सक्रियता के संबंध में भी यही बात होती होगी।
इन विभिन्नताओं के होते हुए भी कुछ कोशिकाद्रव्य इंद्रियकोशिकाएँ (Cytoplasmic organelles) ऐसी हैं जो सभी प्राणियों में और उनकी हर प्रकार की कोशिकाओं में पाई जाती हैं। ये हैं : (१) माइटोकॉण्ड्रिया (Mitochondria) और (२) गोलजी पदार्थ।
कामिल्लो गोलजी इटली का एक तंत्रिकावैज्ञानिक था, जिसने श्वेत उलूक (Barn Owl) की तंत्रकोशिकाओं में एक कोशिकाद्रव्यी इंद्रियकोशिका का पता लगाया, जो उसके नाम से ही विख्यात है। अंडों में माइटोकॉण्ड्रिया और गोलजी पदार्थ अंडपीतनिर्माण में योग देते हैं। स्परमाटिड के स्परमाटोज़ोऑन में परिणत होते समय गोलजी पदार्थ ऐक्रोसोम बनाता है, जिसके द्वारा संसेचन में वह अंडे से जुट जाता है। माइटोकॉण्ड्रिया से स्परमाटिड का नेबेनकेर्न (Nebenkern) बनता है और जिन जंतुओं में मध्यखंड होता है उनमें मध्यखंड का एक बड़ा भाग। ग्रंथि की कोशिकाओं में स्रावी पदार्थ को उत्पन्न और परिपक्व करने में माइटोकॉण्ड्रिया और गोलजी पदार्थ सम्मिलित हैं।
इलेक्ट्रान सूक्ष्मदर्शी से फोटो लेने पर पता चलता है कि प्रत्येक माइटोकॉण्ड्रिया बाहर से एक दोहरी झिल्ली से घिरा होता है और उसके भीतर कई झिल्लियाँ होती हैं जो एक ओर से दूसरी ओर तक पहुँचती हैं या अधूरी ही रह जाती हैं। गोलजी पदार्थ में कुछ धानियाँ (Vacuoles) होती हैं, जो झिल्लीमय लैमेला (membranous lamellae) से कुछ कुछ घिरी होती हैं। इनसे संबंधित कुछ कणिकाएँ भी होती हैं जो लगभग ४०० आं० के माप की होती हैं। ये सब झिल्लियाँ इलेक्ट्रान सघन होती हैं। कोशिकाद्रव्य स्वयं ही झिल्लियों के तंत्र का बना होता है, परंतु इसमें संदेह नहीं कि इन झिल्लियों के माइटोकॉण्ड्रीय झिल्लियों और गोलजी झिल्लियों में कोई विशेष संबंध नहीं होता। ऐसी कोशिकाद्रव्यीय झिल्लियाँ ग्रंथि की कोशिकाओं में अधिक ध्यानाकर्षी अवस्था में पाई जाती हैं। ये अरगैस्टोप्लाज्म कही जाती हैं। ये झिल्लियाँ दोहरी होती हैं और इनपर जगह जगह छोटी छोटी कणिकाएँ सटी होती हैं।
कुछ विद्वानों की यह भी धारणा है कि गोलजी पदार्थ कोई विशेष ऑरगैनेल (Organelle) नहीं है। यह माइटोकॉण्ड्रिया का ही एक विशेष रूप है अथवा केवल धानी के रूपपरिवर्तन से बनता है अथवा केवल एक कृत्रिम द्रव्य है, इस संबंध में विद्वानों में अब भी मतभेद है। (मुरलीधर श्रीवास्तव)